Pteridofitas
-IntroducciónLa evolución de las plantas vasculares empezó con las pteridofitas las cuales se clasifican en helechos,licopidos y equisetos .Fueron los primeros en colonizar el hábitat terrestre sufriendo varias adaptaciones como una nueva ruta metabólica a partir de la tirosina que conduce a la formación ácidos fenólicos (protección contra los fitofagos y eliminación de efectos dañinos por autooxidación) ,síntesis de lignina para evitar la perdida de agua por el tallo Entre sus principales características encontramos la alternancia de generaciones con una ramificación dicótoma, rara vez monopódica, son raras las formaciones vasculares secundarias, con una cutícula impermeable, se reproducen por gametos masculinos flagelados, implicando la presencia de agua.
-Objetivos
*General
Identificar los tejidos internos en las pteridofitas
*Específicos
Observar los haces vasculares de las pteridofitas en cortes transversales del tallo
Reconocer los esporangios y esporas en el corte transversal de los soros
-Justificación
Teniendo en cuenta que las pterofitas son las predecesoras de las plantas vasculares “modernas”, su estudio y conocimiento permite abrir una ventana hacia el pasado, de esta manera se puede comprender el proceso de especialización que las plantas desarrollaron para colonizar el medio terrestre.
-Marco teórico
* Colorantes
Verde de metilo es un colorante básico progresiva (es decir, que actúan en una solución de colorante por un pequeño periodo de tiempo en los cortes.
En la histología, que es muy útil para cierto color ácidos nucleicos y enclaves citoplasmática, que colores como la seda, el moco y el cartílago.
Tionina Es un valioso colorante cromático a causa de sus propiedades metacromáticas. Es usado como colorante vital
La safranina (también llamada Safranina O o rojo básico 2), es un colorante biológico, de contraste en histología y en citología.
-Los licopodios
 |
| Imagen 1 : Esquema de la organización morfológica del licpodio |
Son el grupo de plantas vasculares o cormofitas más simples, aunque están más adaptados al medio terrestre que sus antecesores. Los licopodios primitivos eran homosporos; esta característica sigue persisitiendo en algunos generos. (S.Nieto, 1999)
Son pequeñas plantas herbáceas, perennes y algunas epifitas. Es característica la ramificación dicótoma de sus tallos y raíces, así como sus hojas pequeñas, uninervias y sésiles. Sobre ciertas hojas del ápice de las ramas aparecen los esporangios, tras la germinación de la espora se forma un gametofito subterráneo en el que se desarrollan los órganos sexuales y la formación del cigoto. La fecundación depende de la presencia de agua ya que los licopodios presentan espermatozoides flagelados que necesitan un medio acuoso para desplazarse. El aspecto de las esporas es exclusivo de este género ya que tienen el contorno subtriangular. Los licopodios arborescentes fueron muy importantes durante el carbonidero formando bosques extensos que influyeron de forma decisiva en la flora y fauna de la zona. Los licopodios actuales son todos herbáceos. (S.Nieto, 1999)
Los licopodios presentan amplia distribución, se pueden encontrar tanto en climas frios como tropicales, aunque abundan más en los últimos. (S.Nieto, 1999)
-Los equisetos
 |
| Imagen 2: Esquema de la morfología del equiseto |
También reciben el nombre de colas de caballo. Son plantas herbáceas de regiones y zonas húmedas, descendientes de formas paleozoicas de porte arbóreo que se integraban en los bosques del carbonífero. Los equisetos actuales son el unció genero superviviente de la subdivisión esfenofitinas que vivían en zonas pantanosas. Los cuerpos muertos de estos equisetos, junto con los de otras plantas, fueron el origen de los depósitos actuales del carbón de hulla. (S.Nieto, 1999)
El esporofito de los equisetos está formado por tallos aéreos articulados con nudos y entrenudos que presentan ramas verticiladas en los nudos, el tallo principal suele presentar el mismo diámetro en toda su longitud. El rizoma es horizontal, subterráneo y ramificado del que surgen raíces delgadas y los tallos aéreos. Pueden no tener capacidad sintetizadora por lo que la fotosíntesis se realiza en el tallo. (S.Nieto, 1999)
Los esporofitos forman una espiga en el extremo de algunas ramas, y cada uno de ellos lleva de 5 a 12 esporangios sésiles. Las esporas llevan enroscadas unas fibras en espiral que reciben el nombre de elateres, que se desarrollan en ambientes secos para facilitar la dispersión. La germinación de las esporas de lugar a gametofitos verdes dioicos de unos dos centímetros de diámetro provistos de clorofila y de rizoides y dotados de arquegonios y anteridios. Los espermatozoides del grupo son multiflagelados. Tras la formación del cigoto, este genera una planta esporofitica que al principio que al principio parasita al gametofito, pero que pronto se independiza formando tallos y raíces. (S.Nieto, 1999)
Los equeisetos presenta depósitos de silce en la epidermis, lo que las hace ásperas y cortantes por lo que fueron utilizados como abrasivo para la limpieza de utensilios de cocina y para pulir metales y otros objetos. (S.Nieto, 1999)
-Los helechos primitivos
Son criptógamas vasculares típicos con raíces, tallos y hojas. Se han encontrado restos fósiles de plantas muy semejantes a los helechos actuales que se consideran precursores de estos y que se conocen con el nombre de prehelechos. (S.Nieto, 1999)
-Los helechos verdaderos
 |
| Imagen 3: Esquema general de la morfología de los helechos |
Los hechos constituyen la clase filicopsidas dentro peteridophitas. Son plantas perennes sin crecimiento secundario de forma que cada año les crecen nuevas frondes erectos. En las latitudes templadas presentan un porte herbáceo con rizomas horizontales muy superficiales desde los que parten la raíces filiformes; su hábitat son los lugares frios y los sotobosques húmedos y umbrosos. En zonas tropicales y subtropicales pueden presentar un porte arbóreo que puede alcanzar los 25 metros con aspecto de palmeras. También pueden encontrarse algunas especies epifitas y especies acuáticas flotantes.
Los bosques de helechos del carbonífero contribuyeron a formar los actuales depósitos de hulla. Los helechos se diferencian del resto de las plantas vasculares sin semilla en la estructura de la hoja, la anatomía del tallo, la localización de los esporangios y el tipo de desarrollo.
Los helechos típicos presentan tallos con medula, y cuando están maduros suelen ser cortos y rizomatosos, aunque en los helechos con porte arborescente son erectos. Como tejido conductor presenta un xilema que solo esta formado por traqueiras los frondes son grandes hojas compuestas cuyo eje central, denominado raquis, es una extensión del peciolo. Las frondes presentan nervaduras, tejido esponjoso fotosintético, una epidermis protectora y estomas. Las frondes jóvenes están llenas de escamas y pelos que se pierden en la madurez. El limbo puede estar divido en pinnas y estas a su vez en pinnulas. En el envés de las frondes existen unas estructuras llamadas soros en los que se encuentran las masas arracimadas de los esporangios que pueden estar protegidos por el indusio. En algunas especies, como el helecho común, se presentan nectarios que secretan un liquido azucarado. (S.Nieto, 1999)
El ciclo vital es el siguiente: en primavera aparecen los esporangios en el envés de las hojas, son estructuras esféricas que se unen al la fronde mediante un pedúnculo. Los esporangios tienen una pared muy fina rodeada por un anillo de células engrosadas, si el tiempo es seco el anillo se contrae desgarrando la pared del esporangio que libera las esporas. Estas esporas, que se han formado por meiosis, caen al suelo y originan gametofitos planos, verdes y fotosintéticos con forma acorazonada la fase gametofita solo puede vivir en un medio húmedo y necesita el agua para que se unan los gametos. Esta exigencia es compensada por los helechos con mecanismos de reproducción asexual por medio de bulbillos vegetativos, raíces adventicias en el ápice de los frondes o por medio de estolones subterráneos. (S.Nieto, 1999)
Si las condiciones son favorables , el gametofito forma una serie de rizoides que se anclan en el suelo y absorben los nutrientes. En la superficie inferior se forman los arquegonios, que son los órganos sexuales femeninos, y los anteridios, órganos sexuales masculinos. Los anterozoides, curvados en espiral y plurifalgelados, fecundan la oosfera a la que llegan nadando atraídos quimiotacticamente por una sutancia que produce la oosfera. Tras su formación, el cigoto se divide rápidamente convirtiéndose en un embrión que permanece dentro del arquegonio al cabo de un tiempo adquiere capacidad fotosintética lo que hace que acelere su desarrollo y pueda independizarse del gametofito cerrando el ciclo. (S.Nieto, 1999)
Debido a la complejidad que supone englobar las diferentes especies de helechos en estos grupos, se utiliza de forma tradicional otra clasificación que agrupa los helechos en tres grandes subclases: eusporangiadas, leptoesprangiadas y hidropterides. (S.Nieto, 1999)
-Tejidos conductores
A nivel microscópico, pata el transporte de sustancias disueltas en el organismo basta con la difusión basal basada en el movimiento térmico de las partículas disueltas. No obstante , la eficacia de la difusión disminuye con el cuadrado de la distancia de difusión. Ya en el interior de las células especialmente grandes, como los pelos radicales, las células internodales de las caraceas etc… , la difusión sola resulta insuficiente y es complementada por una convección suplementaria en forma de corriente plasmática. En los grandes animales y plantas pluricelulares han acabado formándose unos sistemas conductores especiales en los que se mantienen unas corrientes conectivas masivas. Así como en los animales la corriente tiene lugar en los espacios intracelulares , en las plantas superiores se han formado unas células especiales por las que circulan los liquidos. Estas células extrema y terminalmente diferenciadas ( es decir , no pueden presentar un desarrollo posterior) se unen formando haces conductores. En las hojas los haces conductores son visibles a simple vista: son las “venas o nervios foliares”. En las raíces, el tejido conductor esta incluido en el cilindro central. (E.Strasburger, 2003)
En todos los órganos conductores hay dos tipos de tejidos diferentes: en la parte cribosa ( floema, leptoma; gr. Pholios: corteza: leptos: fino, delicado) , unas células vivas auqneu desprovistas de nucleo y con finas paredes no lignificadas, se encargan de conducir a largas distancias comopuestos oganicos; en la parte leñosa ( xilema , hadroma; gr. Xylon : madera : hadros : duro , fuerte ), circula el agua con iones nutrientes inorgánicos desde las zonas radicales de absorción y a través de células tubulares muertas , vacías y provistas de paredes lignificadas , hasta las hojas, donde el agua se desprende de nuevo por gutacion o por transpiración. (E.Strasburger, 2003)
Tanto en el floema como en elxilema, las células son prosenquimaticas y están orientadas longitudinalmente en el haz conductor. Además, estas hileras longitudinales de células forman vías conductoras. (E.Strasburger, 2003)
-Floema
Por su origen evolutivo y por su capacidad de transporte, las células cribosas no son eficaces al máximo. Tienen un lumen estrecho y se unen a las paredes terminales anguloso-oblicuas de las células cribosas sucesivas de las filas. Estas paredes (en caso de que estén en contacto longitudinal con las otras células cribosas, también las paredes laterales) están atravesadas por plasmodesmos engrosados, que aquí se denominan poros cribosos. Estos están agrupados formando campos cribosos, a cuyo aspecto deben el nombre. En muchos angiospermas, este primitivo sistema conductor se ha desarrollado hasta formar un sistema continuo de tubos cribosos constituido por células alargadas de gran diámetro y paredes transversales u oblicuas atravesadas a modo de cribas: son segmentos de los tubos cribosos. En las formas más desarrolladas, el floema, como en los bejucos y las trepadoras, las paredes terminales transversales constan de una placa cribosa única con poros especialmente grandes. (E.Strasburger, 2003)
Las células cribosas y los segmentos de los tubos cribosos contienen protoplastos vivos con poca cantidad de mitocondrios y plastidios que almacenan almidón y proteínas. El núcleo celular y el tonoplasto, dictiosomas y ribosomas desaparecen tempranamente, el citoplasma y el jugo celular se mezclan (una de las pocas excepciones de la regla de compartimentación) El retículo endoplamastico se transforma en el llamado retículo de elementos porosos, compuesto de túbulos ramificados y cisternas lisas y apiladas. Un componente característico de los elementos porosos poco maduros son los túbulos de proteína P (proteína floemática) Los elementos cribosos, al ser células anucleadas y delicadas duran poco , la mayoría de las veces se colapsan al finalizar el periodo vegetativo y, en las plantas vivaces , son sustituidos por otros nuevos. En las monocotiledóneas perennes como las palmeras, pueden vivir también muchos años. Al igual que los plasmodesmos, los poros cribosos pueden obstruirse por acumulación de calosa. En las eucotiledoneas o en algunas monocotiledóneas con tubos cribosos abiertos o con otros tipos de heridas, los tubos cribosos aparecen obstruidos por tapones filamentos proteínicos o fragmentos de plastidios rotos. (E.Strasburger, 2003)
En los angiospermas, cada segmento de tubos cribosos esta flanqueado por unas células anexas o anejas , las cuales son pequeñas, anucleadas y poseen abundantes mitocondrias. Estas células glanduliformes, que están en conexión con los segmentos de los tubos cribosos a través de numerosos plasmodesmos, suplementan el metabolismo de los elementos conductores anucelados. De esta manera se sintetiza la proteína floematica y se almacena rápidamente de los segmentos de los tubos cribosos, donde a continuación se forman los cuerpos PP, muy organizados, que más tarde se dividen de filamentos. La segunda función fundamental de las células anejas es la carga y descarga de los tubos cribosos, también esto se traduce en características especiales; o se forman en las células vecinas plasmodesmos especialmente numerosos (tipo simplasmatico= simplastico, sobre todo en las plantas tropicales y subtropicales), o la superficie de las células se ve engrosada por el laberinto parietal; tipo apoplasmatico= apoplasmatico, en las plantas de zonas templadas o frías). (E.Strasburger, 2003)
En la angiospermas, el complejo segmento de los tubos cribosos/células anexas procede de una célula madre por división desigual. Las células cribosas de las gimnospermas y los helechos carecen de células anexas. Sin embargo, en las plantas existen células parenquimaticas ricas en proteínas que están conectadas a las células cribosas del mismo modo lo que están conectadas a las células cribosas del mismo modo que lo que están las células anexas a los segmentos de los tubos cribosos, aunque, a diferencia de estas, no proceden de una misma célula progenitora. Estas células reciben el nombre de células albuminiferas, células de strasburger. (E.Strasburger, 2003)
-Xilema
la corriente de transpiración se desplaza a través de las células tubulares, cuyos protoplastos mueren al alcanzar su capacidad funcional y desaparecen por autolisis; un impresionante ejemplo de muerte celular programada(apoptasis) en las plantas. Solamente quedan las paredes celulares lignificadas y atravesadas por pateaduras. Hay dos tipos de elementos: (traqueales) que se encargan de la condición del agua; traqueidas y tráqueas (vasos leñosos). Las traqueidas son células de lumen estrecho, alargadas, con paredes terminales angulosas y oblicuas; cubiertas de abundantes punteadas, a través de las cuales están unidas con las traqueidas contiguas en dirección longitudinal. La resistencia a la corriente en las filas de las células traqueidales es relativamente elevada. Es considerablemente menor en los segmentos traqueales, más cortos y de lumen amplio, en los que las paredes terminales están muy perforadas o han desaparecido secundariamente por completo. El diámetro mayor de los vasos leñosos (de 60 a más de 700um) visibles ya a simple vista de forma de (poros de la madera) está relacionado con el hecho de que los segmentos jóvenes de los vasos leñosos crecen con anchura por poliploidizacion de sus núcleos (8-16n) antes de que las paredes celulares pierdan su capacidad para crecer al engrosarse estas secundariamente. (E.Strasburger, 2003)
La lignificación de las paredes de las traqueidas y de los segmentos traqueales impide al aplastamiento de estas células tubulares, en las que domina la hipopresion debido a la intensa transpiración. Cuando se corta un tallo, el aire es aspirado hacia los vasos leñosos. Además, las tráqueas helicoidales se asemejan estructuralmente las tráqueas respiratorias de los insectos. (E.Strasburger, 2003)
Se encuentran vías conductoras especialmente simples en los musgos cuyos tallitos contienen cordones centrales de células alargadas y vacías con paredes engrosadas (hidroides). En las pteridofitos y las gimnospermas predominan las traqueidas, el diámetro de los túbulos es aquí mayor y la resistencia a la circulación de las paredes terminales disminuye gracias a su disposición oblicua y la de sostén se produjo relativamente tarde en la filogenia. Todavía en las gimnospermas, el troco, que sirve de soporte, está formado principalmente por traqueidas, las tráqueas han evolucionado dependientemente; aparecen ya aisladas en los helechos y las gimnospermas, en toda su plenitud después, en las angiospermas. La función conductora les corresponde a ellas, mientras que las de sostén es asumida por un tejido mecánico especial, concretamente unas fibras (libriformes) leñosas. También las angiospermas, y por tanto los planifolios, presentan todavía traqueidas junto a las traqueas y en el desarrollo ontogenético de los haces conductores, se repite la evolución filogenética a grandes rasgos. Los vasos leñosos más desarrollados en cuanto a la eficiencia impulsora el mayor diámetro y han desaparecido todas las paredes transversales en espacios de hasta 10 m de longitud, mientras que, en otros casos, en distancias de unos pocos cm, han permanecido las paredes transversales (posiblemente para evitar el riesgo de una embolia gaseosa). La capacidad especial que presenta el tejido conductor de los bejucos se explica teleonomicamente por el hecho de que estos, como las plantas trepadoras que son, no tienen troncos que las sostengan –(esta función la desempeñan otras plantas, rocas, muros) y a través de sus delgados tallos, deben proveer de agua a todo un follaje que tienen la dimensión de la copa de un árbol. (E.Strasburger, 2003)
-Haces conductores
En las raíces, en los tallos y en las hojas , el tejido conductor aparece concentrado en forma de haces leñosos (fasiculos,lat fasiculi:hacecillos)El tejido conductor propiamente dicho esta flanqueado a menudo por haces de fibras esclerenmaticas y rodeado de endodermis. En los tallos y las hojas, los haces vasculares forman a su vez una red, mientras que cada raíz del cilindró centra posee un solo haz conductor radial, el cual en realidad es un haz colector. A partir de la disposición de floema y xilema en el tallo se pueden distinguir entre haces concéntricos y colaterales. Los haces concéntricos con xilema interno están extendidos en los helechos; los que tienen xilema externo, en los rizomas y los tallos de las monocotiledóneas. El tipo de haz conductor colateral es mucho más frecuente (equisetos ,gimnospermas y angiospermas. En el tallo la porción leñosa esta siempre dirigida hacia dentro; en las hojas lo esta hacia arriba , al estar estas dispuestas horizontalmente. El haz bicolateral, con dos porciones cribosas , es una forma especial : tales haces se hallan. ej: en las solanáceas y las cucurbitáceas. (E.Strasburger, 2003)
Cuando la porción leñosa y la cribosa estan directamente en contacto, se habla entonces de haz conductor cerrado. Está formado completamente por tejido adulto. Este tipo de haz es característico en las monocotiledóneas y tiene importantes consecuencias para el crecimiento de estas plantas. En cambio, la mayoría de los haces en gimnospermas y dicotiledóneas son abiertos, es decir, entre floema y el xilema se intercala una capa de meristema: el cambrium fascicular. Transversalmente resalta la disposición regular de las células, de pared particularmente delgada. Este cambrium desempeña un papel decisivo en el crecimiento secundario del tallo. (E.Strasburger, 2003)
-Análisis de resultados
 |
| Imagen 4: Corte transversal de la hoja del helecho sin aplicar ninguna tincion , se reconoce la agrupación de esporangios denominada soros, objetivo 10 x |
 |
| Imagen 5 : Esporangio del helecho vistas con el objetivo de 40 x , en su interior están las esporas contenidas gracias a la capa mecánica y las células del labio |
 |
| Imagen 6 : Esporangio , objetivo 40 x , presenta una dehiscencia longitudinal , la capa mecánica se contrae y abre los labios liberando las esporas |
 |
| Imagen 7 : corte transversal del tallo de helecho visto con el objetivo 10 x , con la tincion verde de metilo , en el se pueden apreciar varios tipos de celulas |
 |
| Imagen 8: Corte transversal tallo de helecho , objetivo 10 x , con tincion de tionina |
 |
| Imagen 9 : corte transversal del tallo del helecho, objetivo 10 x , con tincion de safranina |
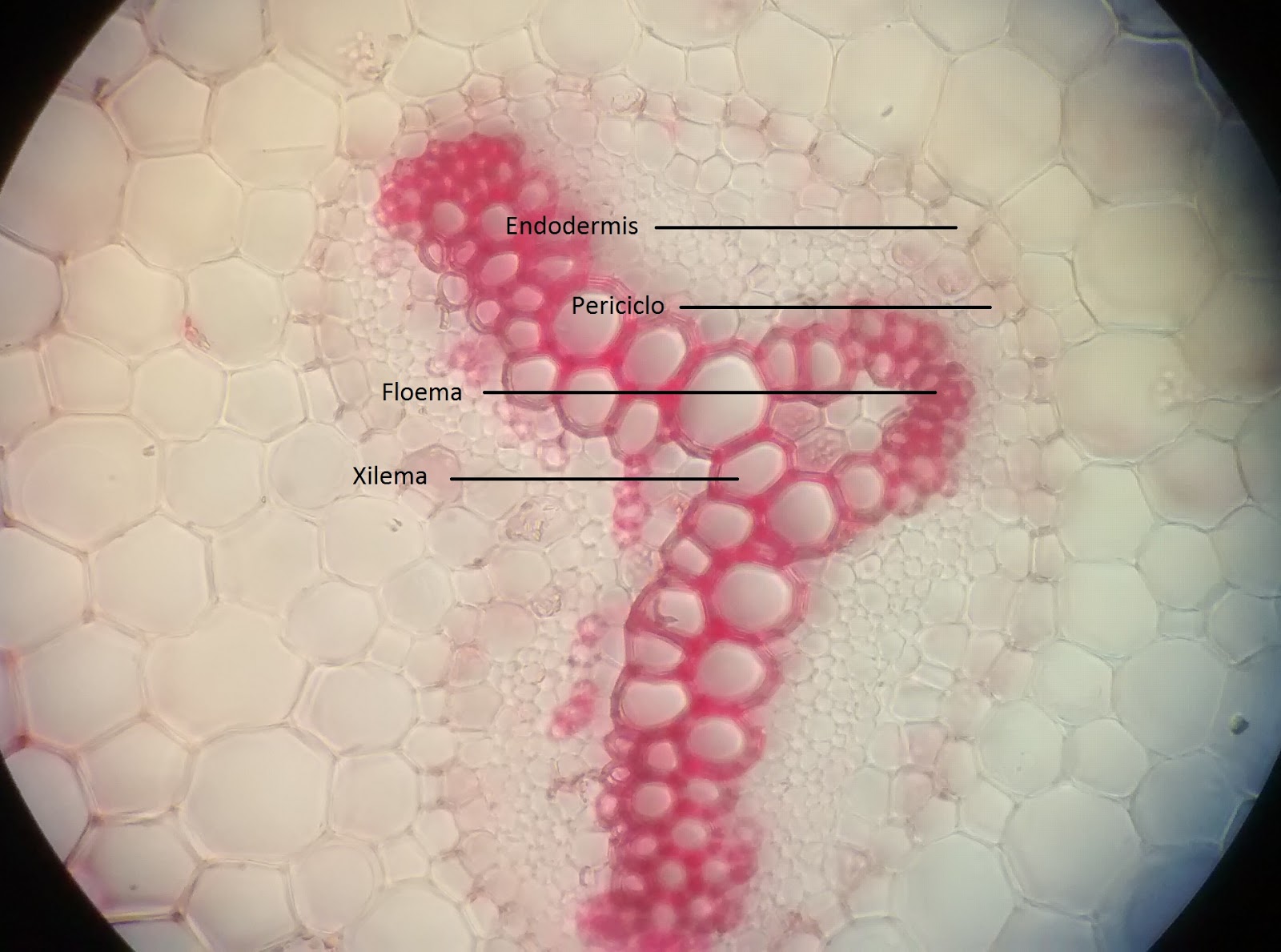 |
| Imagen 10 : corte transversal del tallo de helecho con tincion de safranina visto con el objetivo de 40 x , en el se distinguen las células de xilema , las mas grandes y claras , el floema esta compuesto de celulas pequeñas y se ve de un tono mas definido , también se aprecia la endodermis que es la caopa que rodea los haces conductores |
 |
| Imagen 11: corte transversal del helecho con tincion de verde de metilo ampliado a 40 x |
{kind=link}
 |
| Imagen 12 :Corte transversal del tallo de equiseto visto con el objetivo de 10 x , tincion verde de metilo |
 |
| Imagen 13:Corte transversal del tallo de equiseto visto con el objetivo de 10 x , tincion tionina |
 |
| Imagen 14 : Corte transversal del tallo de equiseto visto con el objetivo de 10 x , tincion safranina |
 |
| Imagen 15 :Corte transversal del tallo de equiseto visto con el objetivo de 40 x , tincion safranina , se aprecian las celulas del floema con un color definido , el xilema no absorve el colorante , vemos la capa de la enpidermis , celulas alargadas en dorma de ladrillo |
 |
| Imagen 16 :Corte transversal de licopodio objetivo 10 x tincion safranina |
 |
| Imagen 17: Corte transversal de licopodio objetivo4 x tincion tionina |
 |
Imagen 18 :Corte transversal de licopodio objetivo 4 x tincion verde metilo   |
Bibliografía
A.Greulach, J. (s.f.). Introduccion a la botanica moderna. Internacional.
E.Strasburger, F. (2003). Tratado de botanica. Ediciones omega
.
M.Holman, W. (s.f.). Botanica general. Uteha.
S.Nieto. (1999). Ciencias de la naturaleza. Rezza.
Elaborado por:
Henrry Paez Mendieta
Jose Alejandro Jimenez Palacios
Hola buenas tardes muy buena publicación la verdad se ve muy a detalle las esporas quería saber si aún conservas la foto original de las esporas pero sin las señalizaciones es que me gustaría utilizarla para una tarea
ResponderBorrar